


で、
ここに出ていたMichael Miller氏および Ulrich Schliewen氏の以下の記事が面白い人には面白いと思うので訳してみました。アピストの遺伝子解析したら、こうでしたよ、って話です。まぁ、あまりニーズはないのはわかっているんですが、欧米の人たちが何をどう考えて分類しているのかを知ってみたい自分には面白かったです。
かなり専門的な内容も含まれていますが、一般向けに解説もついてます。専門でない自分がわかりやすいように意訳もしながら、訳したことだけが気がかりでなりませんが、勘弁してくださいませ。。
The Molecular phylogeny of the genus Apistogramma-
a working hypothesis
アピストグラマ、アピストグラモイディス、タニアカラといった種の系統発生的な関連性を同定するためにKoslowskiは2002年に主に形態および行動の特徴から分類を行うことで、『系統lineages』『グループgroups』『集団complexes』を定義した。
本稿ではこのKoslowskiの分類案を基に我々の分子系統発生研究データの一部を紹介していく。
まず、この分子系統学に不慣れな読者の皆様のためにいくつかの基本的な用語を背景を含めて説明したい。
はじめに全ての遺伝情報がどのひとつの細胞にも含まれていること、”ゲノム”とは個々の器官における全遺伝子座の合計のことであると知って頂きたい。そして、一般に動物においてはその情報の多くは細胞の核にあり”核座”とか”核遺伝子”と呼ばれている。(訳注:核には核のゲノム=”核座”、葉緑体には葉緑体のゲノムってこと。)また、各細胞はミトコンドリアと呼ばれる細胞小器官をもっており、この小器官は細胞にエネルギーを供給しているのだが、このミトコンドリアは独自のゲノムを持っている。
これらすべての遺伝情報はDNAという特殊な分子によって2重らせん構造の中に保持されている。このDNA分子はたった4つの異なるヌクレオシド塩基から出来ており、このたった4つの塩基の並びが複雑な組織の構築、維持、生殖に必要な情報をコードしているのだ。生体ではこの遺伝情報を”読む"ための特殊な酵素が塩基を連続的に読み取ることでタンパク質合成の青写真をつくり、最終的には各パーツとなっていく。
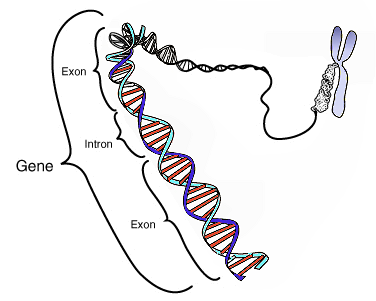
しかしながら、このゲノムはタンパク質へと合成されていく遺伝情報のみをコードしているわけではなく、このコードエリアの中には明らかにタンパク合成をおこなっていない非コードエリアも散在している。
実はダーウィンの示した自然"選択"はこのコード領域と非コード領域に異なった影響を与えるのだ。ここが分子生物学的なプロセスにおいて重要な点で、たとえば、DNA配列の確立的変化である突然変異はコード領域ではコードされるタンパクの機能を維持するよう"選択"されていくため、結果的にコード領域における変異の割合は比較的少ない。対して、非コード領域における突然変異はそのまま"選択"されていっても、結局コードされるわけではないので欠陥タンパク質が作られることはなく有意な影響もない。つまり、非コード領域にはDNAの変異がより高率に蓄積されているのである。
さらに重要な違いが核遺伝子とミトコンドリア遺伝子には存在する。生殖において核遺伝子は両親の物がミックスされる(組み換えされる)が、ミトコンドリア遺伝子は概して母親のコピーである。
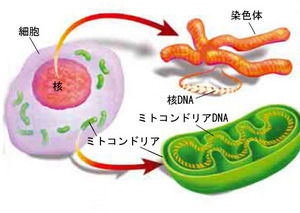
分子生物学では様々な手法に前述の知識が各器官の系統発生的な関連性を明らかにするために用いられている。まず、選択されたコード領域の遺伝子座と選択された非コード領域の遺伝子座のDNA配列を異なるアピストグラマ種の異なる個体たちの解析から決定する。こうして得られた塩基配列の差異の情報を用いて様々な系統学的アルゴリズムで比較、分析を行う。ちなみにこの様々なアルゴリズムというのは様々な進化モデルに即したものであり、進化の連続性につながる生化学的プロセスの確率についての経験的かつ理論的な知識に基づいている。そして、このアルゴリズムは種内における系統発生的な関連性を明らかにすることに長けている。(訳注:具体的に何を意味しているのか自分にはわかりませんw)
今回、我々はアピストグラマ属の系統発生分析において2つの分子データセットを用いた。1つ目は比較的長い配列で全てのアピストグラマグループや近縁種に含まれている核遺伝子、ミトコンドリア遺伝子の双方を含んだ36配列のサブセットからなる。このデータセットはより古代からの系統発生的な関連性を示しうるものである。2つ目はミトコンドリア遺伝子のみのデータでわずか数塩基の短いものである。しかしながら、この短いデータセットは膨大なアピストグラマや近縁種の遺伝子データ用に編集したものであり100を超える種と表現型を網羅している。これらによりおおかたの系統的な関連性が見えてきた。
以下がその結果のまとめである。
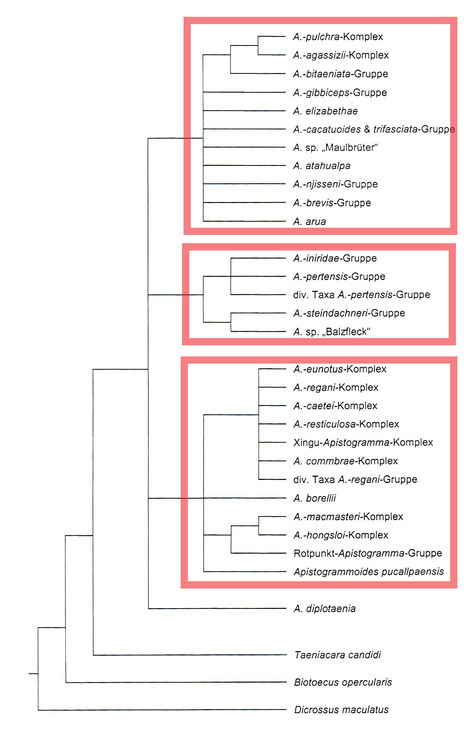
アピストグラマ属(アピストグラモディス属含む)の単系統性が強く示唆された。(最尤法、ベイス推定にて)
2:
アピストグラマ属内における根本的な差異を有意差をもって完全に示すことはできなかった。3つの大きな系統(赤枠)を認めたものの現状その系統内の相互関係についてははっきりしなかった。加えてAp.ディプロタエニアはいずれの3系統とも関与が示されなかったため、4系統にわけた。
3:
この4系統のひとつであるレガニ系は5つのグループにわけることが出来た。アピストグラモイディス-グループ、ボレリー-グループ、ロートプンクト-グループ、マクマステリ-グループ(ホングスロイ-complex、マクマステリー-complexを含む)、そしてレガニ-グループである。このレガニ-グループはさらにコンブラエ-complex、エウノータス-complex(クルジィ-complex、ペバス-complexを含む)、シングーアピスト-complex、レスティクローサ-complex、カエティ-complex、レガニ-complexが認識できる。その他の集団(=complex)の代表個体はいずれの前述の集団にも当てはめることが出来ず、この5グループ内の関連についてもしっかりサポート出来ているわけではない。ただし、ロートプンクト-グループとマクマステリ-グループの姉妹性については言えそうである。
4:
Koslowskiがイニリダエ-グループとスタインダックネリ-グループとともにまとめたペルテンシス-グループはひとつの大きな系統となった。また、未記載種であるAp.sp.”Balzfleck”はスタインダックネリ-グループと姉妹関係にあるとわかった。この系統の中においてペルテンシス-グループとイニリダエ-グループは別グループであり、かつ姉妹関係にあることが認められた。
5:
我々の結果からは、アガシジィ系とトリファスキアータ系は同一の単系統となったが、グループ内の関連や多くの個々の分類群を解決することはできていない。しかし、Ap.アルアはこの系統内で明らかに孤立しており図にみえるようなブレビス-グループと近縁な位置にいるグループではない。
また、カカトゥオイデス-グループはAp.アタフアルパやAp.マウスブルーダー(Ap.バウロウィ)により近縁である。興味深いことにAp.cf.パヤミノニスはニジェッセイ-グループであった。(訳注:このcf.パヤミノニスはA178のことでありその昔マナウス経由で入荷されたタイプと思われる。)
アガシジィ系にはさらに2つの小系が解析できた。一方はギビセプス-グループとAp.エリザベサエがまとめられる系で、もう一方はアガシジィ-グループとビタエニアータ-グループとからなる系である。ビタエニアータ-グループの小系内での位置は明確ではないが、アガシジィ-complexとプルクラ-complexが姉妹関係にあることははっきりした。しかしながら、シングー川流域のある分類群はプルクラ-complexであるとみなされていたが、よりアガシジィ-complexに近縁であると判明した。(訳注:おそらくAp.sp."Blauspiegel" A230のこと)
これらの結果はおおかたKoslowskiの分類を確認するものとなった。また、いくつかの未分類であった表現型をいずれかの系統、グループ、complexに当てはめることは出来たが、多くの物はいまだ明らかにはならなかった。
(注釈:原文のcomplexにあたる言葉は集団だとは思うのですが、なんかしっくりいかなかったのでそのままcomplexで表記しています)
いやぁー、すごいですね~。アピストの分類にここまで頑張ってくる人がいることに感動です。まずは敬意を表したいです。
しかしながら、疑問はつきませんね!
例えば、どういった部分を反映していた塩基を解析に用いたのか?
そもそも解析した塩基が何に関わっていたものかどこまで把握しているのか?
じゃあ、、ディプロのほうが尻ビレの伸条数が多いアピストグラモイディスよりも別種ということか!
たしかに、いろいろ独特だけど。。そこまでなのか・・・とか、
トリファとカカトとアガシジィが同じ系統になってしまうような塩基の選び方は本当に妥当なのか・・・
などなど突っ込みどころはいろいろですが、少なくともある種の定規でアピストの遺伝子を計ってみるとこのように分けられるようです。正しいとか正しくないではなく、この解析ではこうだったということに尽きますね!
参照
Miller, Michael & Ulrich Schliewen. 2005. The molecular phylogeny of the genus Apistogramma ? a working hypothesis.
p. 22-25. in Stawikowski, Rainer (ed.). Sudamerikanische Zwergcichliden / South American Dwarf Cichlids
こんなネタよくやるなぁー、って思ったらクリックぷりーず!

2/01/2015
Tags :
アピスト総論っぽいやつ
Subscribe by Email
Follow Updates Articles from This Blog via Email
12 Comments
長文翻訳ご苦労様でした。
Reply Delete別の尺度でみても同じような結果になるのかが知りたいところですね。
それにしても、系統の枝分かれが少なすぎる・・。
いくつかの種から一斉に放散・分化したのかな?
taketatuさん
Reply Deleteいつもコメントありがとうございます!
たいへん励みになります。
仰れる通り違う物差しではどうなんだろうという疑問が残ります。
いろいろ検討してこの物差しなんでしょうが、十分なものではないようですね。
少なくとも必要なデータを含んではいるともいますが、感覚的には何か大切な要素を含む塩基が足りていないのではなんて素人なりに思ってしまいます。
この辺はなかなかプロにも難しいようで、また遺伝子解析でわかった限界みたいな話も書いてみようと思ってます。
まいどです^ ^
Reply Deleteダメだ?、難しすぎる……orz
理系のはずが、
美形だったか……(逃)
そういえば、
95年のフィッシュマガジンには、
サエさんが、アガシジィグループに
なってたような(汗)
この表を見る限り、
遠からず近からず…
っと、
いったところでしょうか(汗)
僕には、
別種にしか見えません(笑)
ただアピストという種を、
色んな目線で見ることは、
凄く大事だと思います^ ^
僕なんか、
ユルエンシスを見る度に、
「お前は、カナリヤか!!(笑)」
っと、ツッコンでますから(苦笑)
ココナゴさん
Reply Deleteどうもです!
こんなネタにお付き合いいただきありがとです!
個人的には
「お前はカナリヤか!!」のほうが大事な話ですよ、我々には。
それにまちがいなく理系より美形が優遇される世の中ですから、
ココナゴさんがほんと美形でよかったーって話ですね!
自分なんて理系でドイツに流刑ですからw
系統、グループ、コンプレックス(集団)っていう分け方をここの人たちはとりたいみたいですね。それぞれどのレベルの差異かは明記されてませんが一定のルールでわけるとこうなるようですから、そのルールの中ではもちろん正しい分類になっているはずです。
だから、ここから言えるのは見方によっては遺伝子レベルからもエリザベサエはペルテンシス系よりもアガシジィ系ですねってくらいですよw
翻訳お疲れ様です?
Reply Delete結果まとまったのか、まとめる方向で解析したのか?実に興味深い内容です。
細かく枝分かれさせるのが学者さんだと思っていたので?
しかし頑張ったな?趣味でやったのかな?
何気に衝撃的な分類ですよね!エリザベが…
ディプロはそーだろーなーと納得したりして。
けんけんさん
Reply Deleteねぎらっていただきありがたやです!
文章をそのまま信じるならば、Koslowskiの分類を裏付けることを目的の一部としていたと思います。彼らの用意したデータセットでの最大公約数がこれなんじゃないかなー、なんて思ってますが真意は著者のみぞ知りえますw
ディプロはやはり感もありますよねー。
そうなるとディプロはネグロの本家とオリノコのcf.ディプロの2種のみのディプロ系で確立しそうですね。
エリザベはアガシジィやカカト、ビタエニなど共通祖先からギビセプスと一緒に分岐したとの理解になりますね。まぁ、著者も有意な差はなかったといっているのであれですが。。
個人的には解析に用いた塩基数がたりないんだと思います。おそらく予算の問題だと思います。
どうもです~
Reply Deleteマジ翻訳お疲れ様です? (笑)
この感じで近い種は交配出来るのかな?
エリザベとペルテンは無理だわな~(笑)
パパさん
Reply Deleteパパさんまでコメントありがとうございます。
いやぁー、正直途中でやめようと思たんですが、魚がやもめ暮らしなホングスロイしかいないので他に書くことないんですよね。
交配はどうなんでしょうね?
近縁のcomplex同士のほうが感覚的にはF2まで取れるんじゃないかと予測しますが、予測です。まぁ、つまり、わかりませんので実験室で是非w
直接の回答にはなりえませんが、パパさんの御質問に絡むような論文1個知ってるので気力がもてばまたアップしてみます。
まいどです~!超亀コメすみません~m(_ _)m
Reply Deleteおー!例の本ですね(笑)読んでもらえてよかったです(^-^)
ペルテンシスGとスタインダックネリーGがわりかし近縁なのが意外です。
あと、マクマステリーGとエウノータスGはもっと近くていいのにと思いますね~。
そして、やはりディプロは定位置ですな(笑)
AGさん
Reply Deleteまいどです!ぜんぜん亀ではございませんよ!
例の本をここで活用させて頂きました。教えて頂きありがとうございました!いい本ですね~これは。
文中から察するに、エウノータスCとマクマステリGの関連性はわかんないようですよ。遠いのか近いのかも今回の解析ではでないようです。ただ、今回の定規だとレガニグループとマクマスグループという違うグループに計られるようですね。
両者はどうみても近縁に感じますよね~。
スタインダックネリGとペルテンGが近いってのは他にもどこかで見た気がしますが、DATZ本だったかもしれないので同じ元ネタですね。
ディプロにはだれも異論はないのが楽しいですね♪
アピストグラモディスもアピストグラマでいい気がしてきます。
内容については・・・、何か難しくてよく分かりません(汗)
Reply Deleteしかしこの切り口でもっと深く分類していけば面白い結果が出てくるかもしれませんね!
cf.表記がなくなったり、もしかしたらドワシクにアピスト以外の種が登場したり(笑)
個人的には、Ap.cf.パヤミノニスはニジェッセイ-グループとありますが、Ap.パヤミノニスはカカトゥオイデス-グループに属しているのかが気になるところです。
デスモさん
Reply Deleteどうもです!
そうでねー、研究室の予算によると思いますが、少しずつでもやってもらえたら趣味人としては楽しいですね!
内容は自分もちゃんとはわからない部分があるので、その辺は訳が適切でないかもしれません。
Ap.パヤミノニスは2005年の同書ではcf.がそうだったからなのか、なんなのかはわかりませんがニジェッセイグループに入ってます。
本種の標本を解析した結果かどうかまではわかりません。。
個人的にはニジェッセイグループには感じられませんね。カカトやファスカル、アタフアルパとかの方に近縁だと思ってますw